
The aim of this study was to gain a better understanding of how the plant community in vineyards could potentially be manipulated to improve bee diversity.
Global declines in pollinating insects
Wild pollinating insects are declining in abundance and diversity worldwide. This is due to many interacting stressors such as climate change, pathogens, intensive farming practices with high levels of agrochemical inputs, and landscape transformation, which leads to a loss of pollinator habitats.1 More than 75% of global food crops and more than 85% of wild flowering plant species rely on pollination provided by animals, mainly insects. Therefore, the decline in pollinators is of major concern for food security and the continued functioning of natural ecosystems.3
Even though wine grapes do not depend on insect pollination, vineyards that are managed in an ecologically sensitive way can provide pollinator-friendly habitats and, in doing so, support this important group of insects in farming landscapes. Factors that promote them include flowering plants in vineyard cover crops that provide essential food sources such as nectar and pollen,4 reduced disturbance in vineyard inter-rows which promotes ground-nesting pollinators,7 and semi-natural features in the landscape such as fallows and solitary trees which provide nesting habitats.8 The potential contribution of viticultural landscapes to pollinator conservation is significant because conservation in farmland is becoming an increasingly important part of global conservation strategies.3
How do flowering plants in vineyard inter-rows influence wild bees?
A collaborative study during 2019 – 2020 between Stellenbosch University and the University of Natural Resources and Life Sciences, Vienna, assessed how different aspects of the plant community in vineyards influence wild bee diversity.5 Previous work highlighted the importance of floral abundance (total coverage of flowers) in vineyards for bees.6,7 This study followed this work to focus on the influence of flowering plant taxonomic and functional richness on bee diversity.
Hypothetical plant assemblages illustrate the difference between taxonomic and functional richness:



FIGURE 1 – ASSEMBLAGE A. High taxonomic richness (many different species) but low functional richness (they are all ecologically similar and belong to the same plant family).


FIGURE 1 – ASSEMBLAGE B. High taxonomic richness and high functional richness (a wide variety of ecological, morphological, and life-history traits among species).
Taxonomic richness refers to the number of species in an assemblage, whereas functional richness refers to the variety of ecological, morphological, or life-history traits (for example, flower colour, shape, longevity, and flowering season) among species in an assemblage. In the hypothetical plant communities in Figure 1, Assemblages A and B have equal taxonomic richness, but Assemblage B has higher functional richness. Plant and animal communities with high functional diversity support more diverse ecological functions and more diverse responses to disturbance, which improves the functioning and resilience of these communities.10

FIGURE 2. Data collection of plant assemblage in an Austrian vineyard.
We used data that were previously collected from the Stellenbosch region in South Africa and from Carnuntum and Neusiedlersee-Hügelland in Austria (Figure 2). For plants and bees, we selected traits that are relevant to pollination (Figure 3) and then calculated the functional richness of plants and bees for each site based on these traits. Additional factors were considered in the analyses, including vineyard management approach (organic vs non-organic), total vegetation cover in vineyards, amount of semi-natural vegetation in a 500 m radius around the sites, and number of woody structures (woodland patches, tree lines and solitary trees) in the surrounding landscape.
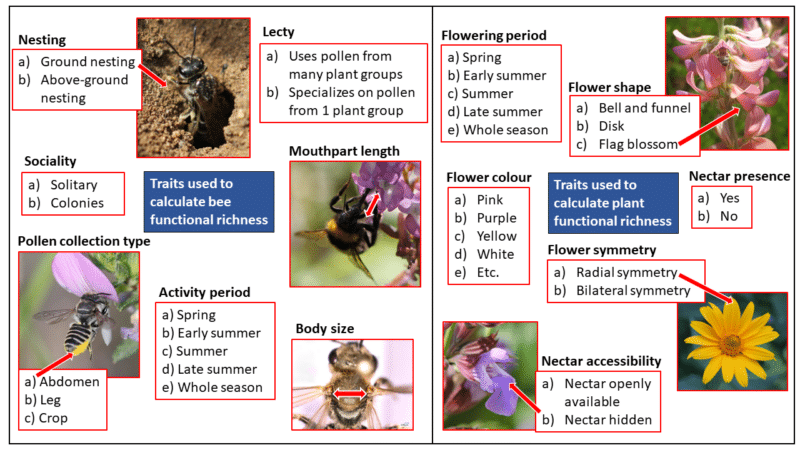
FIGURE 3. Traits used to calculate wild bee and flowering plant functional richness.
The most prominent result was that high flowering plant functional richness promoted wild bee taxonomic and functional richness across management regimes and regions (Figure 3). This means that a wide range of flowering plant species that vary in their traits best promote a wide variety of wild bee species and traits.
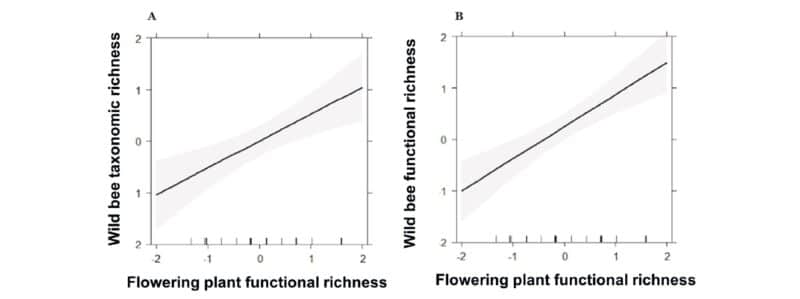
FIGURE 4. Effect plots showing the influence of flowering plant functional richness on a) wild bee taxonomic richness, and b) wild bee functional richness. The shaded area indicates 95% confidence intervals.
This positive relationship may be explained by the fact that individual bee traits were associated with distinct plant traits. For example, in Austrian vineyards, short-tongued bees benefited from yellow flowers with radial symmetry and hidden nectar, likely because their mouthpart morphology allows them to easily access hidden nectar compared to other insects such as flies (Figure 4). This type of trait matching between flowering plants and flower-visiting insects is important for effective pollination of the plant, and for the insect to access its food resource, pollen and/or nectar (Figure 4).2

FIGURE 5. An example of trait matching between pollinators and flowering plants: Short-tongued bees are effective pollinators of radial, yellow flowers with hidden nectar, which often belong to the Asteraceae family.
Management implications
Increasing the functional diversity of flowering plants in and around vineyards promotes wild bee diversity. This could be done by selecting cover crop mixtures with diverse floral traits (e.g., by selecting plants from different plant families) and by reducing tillage that favours a wide variety of plant traits. This work shows potential to fine-tune vineyard practices to enhance pollinator diversity and plant-pollinator networks in viticultural landscapes. It would be supported by further research on how this could be optimised for different conditions in the South African context.
References
- Dicks, L. V., Breeze, T. D., Ngo, H. T., Senapathi, D., An, J., Aizen, M. A., Basu, P., Buchori, D., Galetto, L., Garibaldi, L. A., Gemmill-Herren, B., Howlett, B. G., Imperatriz-Fonseca, V. L., Johnson, S. D., Kovács-Hostyánszki, A., Kwon, Y. J., Lattorff, H. M. G., Lungharwo, T., Seymour, C. L., … Potts, S. G. (2021). A global-scale expert assessment of drivers and risks associated with pollinator decline. Nature Ecology and Evolution, 5(10), 1453–1461. https://doi.org/10.1038/s41559-021-01534-9
- Garibaldi, L. A., Bartomeus, I., Bommarco, R., Klein, A. M., Cunningham, S. A., Aizen, M. A., Boreux, V., Garratt, M. P. D., Carvalheiro, L. G., Kremen, C., Morales, C. L., Schüepp, C., Chacoff, N. P., Freitas, B. M., Gagic, V., Holzschuh, A., Klatt, B. K., Krewenka, K. M., Krishnan, S., … Woyciechowski, M. (2015). Trait matching of flower visitors and crops predicts fruit set better than trait diversity. Journal of Applied Ecology, 52(6), 1436–1444. https://doi.org/10.1111/1365-2664.12530
- (2019). Summary for policymakers of the IPBES global assessment report on biodiversity and ecosystem services. https://doi.org/10.5281/zenodo.3553579
- Kehinde, T., & Samways, M.J. (2014) Insect-flower interactions: network structure in organic versus conventional vineyards. Animal Conservation, 17, 401-409. doi:10.1111/acv.12118.
- Kratschmer, S., Pachinger, B., Gaigher, R., Pryke, J. S., Schalkwyk, J. Van, Samways, M. J., Melin, A., Kehinde, T., Zaller, J. G., & Winter, S. (2021). Enhancing flowering plant functional richness improves wild bee diversity in vineyard inter-rows in different floral kingdoms. Ecology and Evolution, 11, 7927–7945. https://doi.org/10.1002/ece3.7623
- Kratschmer, S., Pachinger, B., Schwantzer, M., Paredes, D., Guernion, M., Burel, F., Nicolai, A., Strauss, P., Bauer, T., Kriechbaum, M., Zaller, J. G., & Winter, S. (2018). Tillage intensity or landscape features: What matters most for wild bee diversity in vineyards? Agriculture, Ecosystems and Environment, 266, 142–152. https://doi.org/10.1016/j.agee.2018.07.018
- Kratschmer, S., Pachinger, B., Schwantzer, M., Paredes, D., Guzmán, G., Goméz, J. A., Entrenas, J. A., Guernion, M., Burel, F., Nicolai, A., Fertil, A., Popescu, D., Macavei, L., Hoble, A., Bunea, C., Kriechbaum, M., Zaller, J. G., & Winter, S. (2019). Response of wild bee diversity, abundance, and functional traits to vineyard inter‐row management intensity and landscape diversity across Europe. Ecology and Evolution, 9(7), 4103–4115.
- Paiola, A., Assandri, G., Brambilla, M., Zottini, M., Pedrini, P., & Nascimbene, J. (2020). Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Science of the Total Environment, 706, 135839. https://doi.org/10.1016/j.scitotenv.2019.135839
- Winter, S., Bauer, T., Strauss, P., Kratschmer, S., Paredes, D., Popescu, D., Landa, B., Guzmán, G., Gómez, J. A., Guernion, M., Zaller, J. G., & Batáry, P. (2018). Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. Journal of Applied Ecology, 55(5), 2484–2495. https://doi.org/10.1111/1365-2664.13124
- Woodcock, B. A., Garratt, M. P. D., Powney, G. D., Shaw, R. F., Osborne, J. L., Soroka, J., Lindström, S. A. M., Stanley, D., Ouvrard, P., Edwards, M. E., Jauker, F., McCracken, M. E., Zou, Y., Potts, S. G., Rundlöf, M., Noriega, J. A., Greenop, A., Smith, H. G., Bommarco, R., Pywell, R. F. (2019). Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield. Nature Communications, 10(1). https://doi.org/10.1038/s41467-019-09393-6
For more information, contact René Gaigher at reneg@sun.ac.za.
Click here to get your copy of WineLand Magazine and here to subscribe to our newsletter.













